Essays and Theories
![]()
Essays and Theories
A new car may run 70 or 80 thousand miles without major repairs, but then the carburetor or the transmission will go, and soon progressively larger systems become vulnerable to failure. You might think that this is a result of good engineering: the designers have thought carefully about the car's major parts and decided how much to invest in durability of each one, balancing initial cost against the later cost of repair. The lifespan of the car as a whole creates a context for this calculus: if the car is likely to be driven a maximum of 150 thousand miles, and if a particular part already has an expected life of 300 thousand miles, then it would not be cost effective to spend even one more penny to enhance the durability of that piece.
Perhaps it is this sort of cost-conscious engineering that drives the designers in Detroit, or perhaps there's an element of planned obsolescence in their thinking. Does the sales department meet with the engineers to arrange for the cars to be durable, but not too durable? Are some crucial parts designed to wear out even though it would be no more expensive to make them last longer? Absent a deep faith in corporate morality, we must entertain the possibility that machinations of this sort could be part of the industry's workings.
Even if we're cynical about corporate America, we may still harbor a deep faith in Nature. Our bodies were designed by Evolution, meticulously over a half billion years. Evolution may not be kind or even smart; but she certainly has the motivation to make a durable product. The entire world of experience has been integrated into the human blueprint. If our fingers are nimble and our minds agile, if we can adapt and learn, and sense danger with pain and sexual pleasure with great joy, then we have Evolution to thank for all this bounty. And if we get old and die - certainly Evolution was doing her best to keep us strong and healthy. There must have been factors beyond her control...
So it may come as a rude shock to realize that planned obsolescence is part of Evolution's way. Our most cynical suspicions about Detroit pale beside the reality of what Evolution has done to us: Our bodies are designed with malice aforethought to deteriorate with time, to decay and to die. Aging and death are not incidental failings, or a piece of unfinished business. We don't just wear out like an old machine. We are designed to fail after a time, with the same deliberation and the same superb engineering that we recognize in the strength of our muscles or the resilience of our livers. Evolution has specified our mortality just as surely as she has designed us to eat and to breathe, to work and to reproduce.
This is a conclusion from which we recoil. Nature's sweet name is spoken with reverence and gratitude. For some of us, Evolution is the closest thing we have to a God. Suspicions of her motives do not sit easily with us, and we resist mightily the notion that it is she and none other who has slipped us the Black Spot. Even researchers in evolutionary biology have been slow to recognize the signs of purposefulness in the cycle of birth, growth, reproduction, decline and death which we see all about us. But signs there are everywhere, once our eyes are opened to them. The one that brought the issue home to me most vividly is the story of dietary restriction and life extension. In experimental studies, animals that are fed less live dramatically longer. If Evolution knows how to help an organism to dodge senescence when food is scarce, why has she withheld this blessing when resources are abundant?
The first experiments in which mice fed on a near-starvation regimen lived substantially longer than their full-bodied, contented brothers were published in the 1930's. There was a steady trickle of follow-up, but for 50 years these reports remained in a backwater, failing to generate much interest even within the esoteric field of senescence biology. Perhaps the irresistible force, in the form of man's quest for the fountain of youth, had come upon the immovable barrier, in the form of his love of food.
A few researchers continued these experiments. Variations were tried with different animals, different schedules, different foods. Protein was separated from fat was separated from carbohydrate. It was found that what works best is a nutrient-rich diet, a high-protein diet with all essential vitamins and minerals but reduced calories. Beyond the minimum quantities necessary for good nutrition, it didn't seem to matter much if the diet was weighted toward carbohydrates or protein or fats; frequency of feeding and exercise (that can decrease body weight) were discounted as causes; total calorie count was the one determining factor.
And what's more, the notion of "reduced calorie" seemed to be quite subjective - subjective from the animals' point of view, that is. The important thing was to eat smaller quantities than was the animal's natural bent. Ad libitum feeding is the code word in the literature for the bowl in the cage that is never permitted to empty. The animal eats as much as he wants, whenever he wants. But mice, like humans, are subject to great individual variation. Under ad libitum feeding conditions there are fat mice and lean, and all grades in between. The funny thing about the dietary restriction (DR) effect was that it didn't seem to depend on the absolute quantities of food the animals were eating so much as the fact that they were eating less than they wanted to. Animals that were innately stout could be brought down to normal weight, and they would enjoy even more life extension, statistically, than their normal-size cousins for whom dietary restriction made them quite scrawny.
DR seems to promote health in general, and also to stretch out every stage of the life cycle. In this latter effect, DR is unique, the only potion that science has to offer a latter-day Ponce de Leon. Life has been prolonged by hygiene, nutrition and medical advances, but none of these can be said truly to affect the rate of aging.
Remarkable advances in human longevity have been achieved in the twentieth century. Many of the infectious diseases that used to strike us down in our prime have become rarities in the developed world, and steady progress has been made even with the diseases of old age: heart attack, cancer and stroke. The average life expectancy of a person born in 1870 was 41 years; a century later that figure had grown to 71. This effect is dramatically visible in the actuarial curves, which show how many people are left surviving at each age, from an original cohort born in the same year. The decline in the curve gets less and less steep as the century progresses.
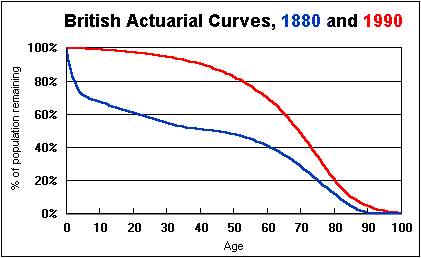
But it's also true that if you look at the right-hand edge of the curve, there's a steep drop off that has resisted medical technology more stubbornly. The oldest survivors in 1880 were 105 or 110 years old; and today, the longest-lived among us survive for about the same 110 years. Most of the century's progress in longevity was achieved without extending the maximum lifespan, but by reducing premature deaths, seeing more and more people through to old age. Only in the last 30 years have the survival prospects for people over 70 begun to advance, and the maximum human lifespan has yet to respond significantly.
The intriguing thing about experiments in dietary restriction is that DR affects the maximum as well as the average lifespan. The oldest mice fed ad libitum live for three years; the oldest DR mice live to age four and a half. Spiders live to a maximum of 100 days if they eat freely, but the oldest DR spiders live 140 days. The appearance of the animals, their activity level and their resistance to disease all seem to remain better, longer on the reduced-calorie regimen. The effect is not subtle: the great majority of DR mice are still healthy and active and to all appearances they are in the prime of life at an age when the last of the ad libitum animals has died. On this ground, experimenters base their claim that DR actually slows the aging process.
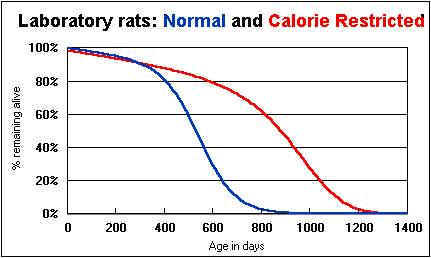
Another striking result: DR animals live healthier as well as longer lives. They are more active and energetic (though they have less calories to burn!). They resist infections and disease more effectively. Their very appearance suggests that they are healthier and more robust than other animals at the same age.
All these conclusions seem to hold for a great variety of living forms. They have been tested with animals as simple as starfish and fruit flies; recent results show the effect for rhesus monkeys. Experimenters haven't been willing to lock up humans and underfeed them, but there are intriguing historical and cross-cultural field studies suggesting that people are no exception.
(This doesn't necessarily mean that people fed an optimal diet - nutrient-rich and restricted in calories - will set dramatic new records for longevity. It may be, for example, that the longest-lived humans at present are already eating significantly less than the rest of us.)
The details of this story makes a powerful case for Death By Design. If the body shows itself capable of living a longer, healthier life under conditions of near-starvation, you have to think that under normal conditions it's holding back. This is the key point: If the DR effect is to be explained on metabolic grounds alone, then "normal" intake of food must be bad for us. Why, then do we get so hungry? And why is it that genetically obese mice who are forced to curb their appetites enough to eat just average quantities of food (and, consequently, to weigh just what average mice weigh) - why do these mice live so much longer than genetically average mice, who under ad libitum conditions are eating the same amount and who weigh the same?
Only in a context where aging is considered a purposeful adaptation do these results make sense. If aging and death are there because they serve the evolving community, then that purpose may easily be overridden by other, greater exigencies. In times of scarcity, evolution has decided that it is advantageous for the individual to prolong survival and to delay breeding until conditions are better for raising children. The individual's hunger is a signal to the effect that the community is in danger, and extinction may be a very real possibility. The survival prospects of every individual must then be maximized. In prosperous times, Evolution may think about the long-term future of the species, and lifespan are limited to achieve an optimal turnover in the population. But short-term thinking is appropriate in a crisis, and enhancing the survival prospects of every individual becomes relatively more important when starvation looms. In normal times, aging and death proceed on a schedule that represents evolution's compromise between the near-term good of the individual and the long-term adaptability of the species; but hunger is a marker of hard times for the species, and the compromise then shifts toward greater survivability and longer lifespan.
The story of dietary restriction and its effect on longevity is one important clue pointing to the conclusion that death is part of evolution's design. Another strand of biological thinking that leads in the same direction is the discovery in the 1970's and 80's of the molecular biology of telomeres. These are biological replication counters that can kill a cell line after it has multiplied a fixed number of times.
Telomeres are tails on the end of every strand of DNA. The replication process that takes place whenever a cell divides always leaves out a stretch of the tail, and the DNA strand becomes shorter with each time that the cell reproduces. The telomere is a buffer zone, a stretch of nonsense DNA that is completely expendable, so for awhile the shortening has no effect on the daughter cell's viability. But eventually the telomere is used up, and the cell can no longer replicate itself.
The curious thing about this process is that it appears to be entirely avoidable. All living things have the capacity, under the right circumstances, to produce an enzyme called telomerase. In the presence of telomerase, the entire DNA molecule is faithfully replicated, and the tail doesn't lose any length. This capability is absolutely essential to the long-term continuity of life. It must be that from egg to hen to egg, the full complement of DNA is reproduced, or the chickens of the world would have long ago disappeared. But outside our gonads, all cells in our bodies have a lifetime limit on the number of times they can reproduce themselves. For skin cells that replenish weekly, this limit is rather long; for brain cells that develop once and last a lifetime, the telomeres are much shorter. Limits on cell division may be essentially related to the aging process, or they may be just a vestige of aging in lower species, but there is little doubt that their operation is purposeful.
Even in one-celled animals, telomeres effect a kind of aging process. Paramecia can reproduce by simple division a few hundred times before they run out of telomere, and the process grinds to a halt. Some time before that happens, the paramecium must find another of its species and conjugate. This process is a primitive forerunner of sexual reproduction: the two organisms come together temporarily, and the DNA in their nuclei is allowed to mix. Then they separate, and each paramecium leaves with some of his own DNA and some of his partner's. And by the way - the telomeres are restored to their full and original length in this process, so each organism is set for another several hundred rounds of asexual reproduction.
The biochemistry of telomeres conveys a distinct impression that this process is a purposeful mechanism and not an accident or side-effect. Telomeres carry no genetic information, and their chemistry doesn't seem to be related to the chemistry of replication. Still, the cell refuses to replicate when the telomere becomes too short. In one-celled creatures, the length of the telomere is nothing but a counter that keeps track of the number of times the cell has divided since its last conjugation. That makes it a measure of how successful a cell line is; the only conceivable purpose is to compel successful cells to share their genes, on penalty of death. Telomeres in one-celled creatures are the model from which aging and death in higher organisms have developed. The lesson is that the rule of the fittest must not be absolute: Evolution wants her most successful children to thrive and to flourish, but not to dominate.
The sharing of genes is a vital part of evolution's plan. The ability to mix and match genes with different, independent adaptive benefits that may have evolved in different individuals is a powerful instrument for evolution's toolbox. Without it, you can imagine how difficult evolutionary progress would be: if an advantageous new gene happened to appear in an organism that was flawed because of some unrelated genetic defect, then the only two choices available would be to accept the new gene and enshrine the flaw in all future generations, or throw away the promise of the new gene in order to cleanse the flaw from the gene pool. But with sexual reproduction (or conjugation, if you happen to be a paramecium) a multitude of new genes can be tested simultaneously by an entire population of individuals. Subsequently, the best genes survive, and eventually come together in individuals who become the super-competitors of their generation. The population is like a million testing labs, working in parallel and combining their results in the end to maximize the benefit. The efficiency of this process is far beyond what could ever be achieved by pure competition of individual specimens.
At any given time, form A may be more adaptive than form B, and it is good that A should flourish. But environmental conditions are subject to change, and it is in Evolution's interest not to discard B entirely. A population's genetic diversity is a hedge against an unknown future. Holding the A gene back so that B can be retained in a small fraction of the gene pool has a short-term cost, but it is good insurance for two reasons: The climate may shift in a direction that makes B once again more viable than A; and there's also the possibility that elsewhere in the population, gene C is emerging, and C in combination with B may just make a better package than C with A.
Just as Evolution has used telomeres to dull the edge of selection's blade, assuring that no one cell-line of paramecia can be so successful that the population loses all its genetic diversity, for this same reason do aging and death exist in populations of higher animals: the species is asserting its prerogative over the strongest individuals' ability to dominate the population to the exclusion of competing forms.
Beyond telomeres and dietary restriction, there are other clues as well that Evolution has freely chosen aging and death. The life cycle of the Pacific salmon is organized around a single breeding event. Just before laying eggs, the salmon is five years old and in the prime of life; she swims many miles upstream, breaches waterfalls by jumping from rock to rock, and displays an agility and determination that defines the phrase "a force of nature". But immediately after breeding, her body undergoes a transformation that looks like aging in time-lapse photography. In a few days' time, the muscles weaken, the skin develops lesions, and the body falls easy prey to all sorts of common ailments. Multiple symptoms tell you that this is indeed an aging process, but its dramatic rapidity tells you that it is no accident.
Another clue comes from the fact that, as we age, everything seems to go at once. Humans are typical in this regard: in old age we lose our eyesight and our hearing; our bones become brittle; our immune bodies can no longer fight common bacteria like pneumonia and chicken pox (shingles); the cardiovascular system, the brain, the ability to regenerate and to heal - we lose all these faculties quite precipitously in the eighth or ninth decade of life. If you think back to the engineer who is trying to balance cost-consciousness against durability, it makes you wonder if all these systems happen to "cost" the same thing, so that it pays to make them comparably durable. Or, more credibly, are we designed as the salmon, with a switch that turns on our demise all at once, in order to assure that no one escapes death.
One last clue comes from sharks and shellfish. Although aging and death do seem to be a near-universal feature of the natural world, there are some creatures that seem to age so slowly that aging plays no real part in their natural life cycles; that is to say, they will almost certainly starve or be consumed by a predator before they begin to feel the ravages of age. If Evolution had been doing her best to suppress aging, though it's a tough challenge for her, you might expect that it would be the newest and most advanced life forms that are the most successful at resisting decline with age; in fact, the opposite is true. Sharks and turtles and oysters and alligators have in common that they have existed in almost the same form for hundreds of millions of years, and that aging is just not an issue in their natural life cycles. This fits well with the theory of aging as Evolution's device for promoting adaptive change: Here is a class of creatures that don't seem to need it; change has proved unnecessary, so aging has lost its raison d'etre, and it has all but vanished from these animals' life cycles.
All these observations support the idea that aging is a purposeful evolutionary adaptation. All creatures want to grow, to thrive, to reproduce - and to die. Our day arrives; we have our chance to compete, to show our stuff; then our time is done, and the torch is passed to our children to continue in our stead. This is how evolution has programmed us, and this is the way all living systems have been designed. To every thing there is a season.
Now at some point I must confess that the scientific themes I have presented here are not yet accepted by the experts in the field. These ideas are respectable science, but at present they are badly out of favor. Most evolutionary biologists will tell you that Evolution cares first and foremost how quickly and how successfully an organism can replicate its genes in the next generation. It's not at all clear to them how this imperative could be overridden by something as subtle as the long-term need for diversity in the population gene pool. The genes within an individual may see many things in their peripheral vision, but what they perceive front and center - what affects most strongly their ability to spread and gain currency - is the ability of the organism bearing them to survive and to reproduce. Evolution operates in a way that is essentially nearsighted. Since aging and death spell no good for the individual, Death by Design is never seriously considered.
With Nearsighted Evolution, how are aging and death to be explained? The answer is more tortured and convoluted than you might imagine. This is because the simple answer - the answer you've probably imagined all your life, when you take time to imagine such things - the most obvious and most ancient answer doesn't work. This is the theory that Things Wear Out.
Knives get dull - why shouldn't our teeth? Wheels get rusty and squeak when they're turned - why shouldn't our joints? Even the stone faces of the ancient mountains are worn and pitted from aeons of exposure to the elements - isn't that just what happens to our skin as we age?
It's a theory with a great deal of intuitive appeal. Thermodynamics has its famous Second Law, (perhaps the most misunderstood scientific principle of all time), saying that all order is in a one-way march toward disorder as the entropy of every closed system increases inexorably over time. This is indeed related to the reason why the knife gets dull and the bearings are subject to rust. But the word we don't notice in the Second Law are "closed system". A closed system is isolated, not interacting with its environment. Certainly that's not true of the knife and the wheel - it's the air and the moisture in the air that rust the wheel; it's the friction of the knife against the cutting board that dulls the blade. For the knife and the wheel, this doesn't seem to matter. The entropy rises in the tool even as the entropy rises in the environment, and the interaction just serves to accelerate the process for both halves of the system.
But living things are living precisely because they have learned to perform a wonderful feat, an end-run on the Second Law. A living organism extracts energy from its environment, and thereby is able to maintain - even to increase - order within itself, while dumping its entropy out into the environment. The total entropy, organism plus environment, is constrained to increase; but as long as it can take in food and unload its waste, the living organism performs a magic that non-living things cannot match: it can grow, it can repair, it can even build itself from scratch, starting with a DNA-blueprint and necessary nutrients.
Every living metabolism includes highly-developed machinery for repair and maintenance. You know that when you break a leg, the bone grows back together. It's less obvious that the DNA in your cells is constantly being checked for errors: specialized molecules creep along the length of the DNA strand, searching for pieces out of place and repairing them on the spot. You can recover completely from a cut in your finger, but if the finger is severed completely, you'll never grow a new one; this limitation is standard in mammals. But consider the starfish: if the starfish loses an arm, not only does the starfish grow another arm, the arm will grow a whole new starfish.
Imagine how complete and how robust is the starfish's system of regeneration - then consider that the starfish ages and dies, with a lifetime of about eight years. Now the inadequacy of Things Wear Out becomes more apparent. Evolution has devised for all of us a system of maintenance and repair that is remarkably efficient. (For the starfish, you would have to say that the system is essentially complete.) In this context, aging and death don't seem at all like an expression of universal physical law. It's a genuine curiosity that Evolution, after creating such intricate and comprehensive systems of repair, has abandoned the job at a stage of completeness that is near perfect. This is the classical problem of aging, the problem that has made generations of evolutionary theorists squirm.
In the 1890's - ever-so-early in the history of two sciences, thermodynamics and evolution, which his genius brought together - August Weismann realized that there was no thermodynamic necessity for the body to degrade over time. If the oak could grow from an acorn, if the starfish could regenerate a severed arm, then Science needed a reason beyond Things Wear Out for the aging process that holds all living beings in its grasp. In fact, Weismann was the first to speculate that the evolutionary reason for aging and death has something to do with the good of the species. He called it "making room" in the environment. When their lease was up, the Old would have the good grace to pack their bags and vacate the premises, leaving space for the Young.
In my minority view, this was an essentially correct explanation, but it must have seemed strange at the time - even to Weismann, who later disavowed it. It has never gained much respect among specialists in the sub-field Evolution of Aging, for the reason we have cited: that it requires a farsighted model for Evolution. Two thirds of the way through the twentieth century, the debate about whether Evolution was nearsighted or farsighted was won by the other side; Evolution had been declared to work on individuals and on their genes, but not on entire populations.
For the (majority) faithful followers of Nearsighted Evolution, aging and death are a great embarrassment. It is plain as day that aging and death are no friend of the individual's capacity to thrive and produce offspring. You can't pull the wool over evolution's eyes. If the nearest, most direct effects of the genes for aging are to limit their own capacity to reproduce and to spread, how can any Darwinian scientist call them "adaptive"? Backed into a corner, the Nearsighted majority has been forced to return to a kind of argument from necessity. The theory that has come to be accepted in the last forty years is based on the Law of Unintended Consequences. Here's how it works:
Individual evolution is always accumulating genes that can augment the organism's capacity to thrive and to reproduce, as early and often as possible. Among possible strategies for achieving these ends, there are those that overheat the body's engines: early in life, when reproduction contributes most powerfully to exponential population growth, the knobs are set to full throttle. The turbochargers and the afterburners that operate at this time promote the individual's ticket in evolution's lottery by getting more offspring out earlier, but (here's the Law of Unintended Consequences) their aggressive and single-minded pursuit of that end is bound to have side-effects. Aging and death are the consequence that we see.
This theory may not seem very compelling on its face. Its premises seem too particular to be an adequate explanation for so universal a phenomenon as mortality. All biological processes have side effects, to be sure - control and regulation of these is one reason that living metabolisms are as complex as they are. Appeal to the Law of Unintended Consequences does not begin to explain why Evolution has failed to provide us the ability to right ourselves once the unintended consequences have befallen us. The theory also fails to account for the DR phenomenon, and the longevity of ancient life forms. It may not seem compelling, yet it has gained ascendancy for lack of competition. Remember that the wear-and-tear argument was debunked early, and all consideration of the good of the species is enjoined by a deep belief in Nearsighted Evolution.
Death by Design is a stronger theory than the Law of Unintended Consequences: It fits comfortably with the story of the salmon and the story of the paramecium, the story of the starfish and the story of the shark. It is the only way to explain the power of dietary restriction to extend life. But if the Law of Unintended Consequences yields to Death by Design, then Nearsighted Evolution will have to fall as well. This will be a small revolution in biologists' perspective, and for this reason it has to be considered a long shot.
My money is on Death by Design, and I don't apologize for Farsighted Evolution. I believe that, looking around us, we see other signs that evolution has managed to take a broader view of what "fittest" means than the consensus of biologists is willing to grant. I'll readily admit that we don't yet understand how Evolution has acquired her wide-field binoculars, or how the optics inside can function as it does. But I submit that Evolution's ability to do this is apparent all around us, in the form of the eyes and wings and complex structures that seem to have evolved of a whole, like Athena springing fully-armored from the head of Zeus. The parts of a wing, or the pieces of an optical cortex in the brain are adaptations that have no selective value until they are fully-formed. How, then, was Evolution guided along the steps necessary to build these complex structures?
Consider, too, the social behaviors that are programmed so intricately into many kinds of animals, from wasps to elephants. Does it seem possible that these could have evolved in a world where natural selection acts only on individuals? Evolutionary biologists recognize a thousand small mysteries like this; I suggest that perhaps they are all part of the same mystery, the mystery of Evolution's binoculars.
True believers are hard at work devising explanations from individual selection for these adaptations that look to all the world like group selection. Evolutionary biologists, like most other specialists, can become so wrapped-up in the systems of thought that they have invented that they fail to notice how tortured their logic may sound to an outsider. But there is one example that has caused the take-no-prisoners brand of Nearsighted Evolutionists to throw up their hands in wonderment at Nature's inscrutability, and that is the evolution of sex.
Sexual reproduction is about as universal as aging. Animals from ants to zebras all reproduce sexually. Plants have flowers and cones. As we have seen, even among one-celled organisms, there is a kind of gene-sharing that serves the function of sex. And yet, the advantages of sexual reproduction accrue solely to the larger population unit; from the individual's point of view, sexual reproduction can't compete with the efficiency of cloning. Reproducing asexually, the individual transmits all its genes to the next generation without help; reproducing sexually means searching for, then selecting a partner and satisfying the partner's own selection criteria. Mating rituals, physical combat and injury are all elements of the competition for sexual partners. When all is done, individuals who succeed in the process transmit only half their genes to each offspring. On the other hand, sexual reproduction offers a major advantage to the species as a whole: All the adaptive benefits gained by different population members can be shuffled, mixed and recombined to create new and fitter individuals. Evolution proceeds far more rapidly with sexual reproduction than it ever could without it.
To the faithful adherent of Nearsighted Evolution, sexual reproduction is the biggest mystery of all. One researcher calls it "the masterpiece of nature", and has written a book by that title, seeking in vain for an explanation in terms of individual selection. For those of us outsiders who were never trained in the prejudice against group selection, it may be difficult to empathize with their wonderment. Group selection seems to us to be visible everywhere, a hallmark of Evolution's farsightedness. It is not just the selfish gene or even the selfish individuals who are the leading runners in Evolution's marathon. Most favored has been the robust community that can adapt and thrive and continue to evolve as circumstances change, and as competing communities respond. Not just genes, not just organisms, but whole ecologies have evolved. Evolution herself has been shaped by the forces of natural selection. Sexual reproduction and the aging process are two qualities whose benefit really accrues to the process of Evolution herself, and the reason for their existence can only be understood in the context of the evolution of Evolution.
-------------------------------------------
If we come to realize that Evolution cares not just about individuals but about communities, if we deeply absorb the message that Evolution has designed our bodies to degrade with age and to die, martyrs all to the communal cause, how does that affect the way we think about ourselves and our relationship to Nature?
For one thing, we will take our herbal medicine and our organic kumquats with a grain of salt. We may never have thought about it in this way, but at root the appeal of the "natural" comes from faith in Evolution. What is natural is part of the environment in which man and his ancestors evolved; hence it is presumed well-suited to us, and we to it. How many articles of grandmotherly wisdom are rooted in this implicit appeal to Evolution - from "trust your instincts" to "eat whole grains"? How many products are marketed by association with the word "natural"? Not just foods, but medicines and herbs, hair and skin ointments, natural fibers, natural furnishings, natural exercise programs, aids to learning and posture supports.
Now we have a reason (beyond our "natural" aversion to commercial hype) to be wary of all such appeals. Sometimes Nature wants what's best for us individually, but sometimes she conspires with out bodies to bump us off - all for the common good, of course. If you're a non-smoker, eating less may be the most effective step you can take to enhance your health and extend your life; but you'll seek in vain for the instinct that supports your willpower when you're denying the body's appetite. Evolution has designed us to enjoy food so that we will put away a comfortable layer of fat for the lean winter which may ever be just around the corner. Nature wants us to eat when there's food to be eaten, but nature wants us to die after a "natural" life span. If you or I have our own ideas about how long we want to live, we may have to play some very un-natural tricks, or even to do battle with nature in order to get there.
Be assured, new and very effective therapies to slow the aging process will be discovered in coming years. The basic biochemical understanding that can make this possible is only now becoming available. My guess is that caloric restriction is the first in a long succession of technologies for augmenting health by tricking our bodies' aging function. No one discovery will conquer aging, because the physiology of aging consists in so many independent processes. (Farsighted Evolution has been working since the dawn of life to hold in check Nearsighted Evolution's relentless quest for longer lifespan, and the result that we experience is multiple, redundant modes of senescence.) But we have every reason to think that each of these separately is amenable to medical intervention, and that ever longer and healthier human lifespan will continue into the foreseeable future.
Technologies for life extension will be eagerly sought by individuals, but it is a legitimate question whether the aggregate effects of longer lifespan will benefit humankind, in a world whose greatest conflicts are associated with overpopulation. I am confident that the global benefits will outweigh the added challenge to population control. The medical assault on aging will not be adding years of infirmity at life's end, but healthy and productive years to the prime of life. As the global economic engine continues to grow in complexity, so is the knowledge required to be an effective citizen increasing and education expanding in importance, not only for youth but through all stages of life. A world of more experienced citizens, in which there is more time for each of us to accumulate knowledge and wisdom, this is likely to be a better world for us all.
The story of Evolution and aging offers another moral, yet more hopeful and uplifting than this one. The economic culture of our generation has embraced unbridled individual competition; for a few, this has unleashed a restless quest for the accumulation of wealth, while the many suffer a disquieting economic decline. Perhaps we could learn from Evolution's insistence on tempering pure competition with universal mortality that diversity of the population is essential to the long-term health of a community. Perhaps the message of Farsighted Evolution can help us to strip away the illusion that our life's purpose is individual achievement, and restore to us a sense of our common destiny. The cycle of life and death can only have meaning in the context of a great evolutionary progression that will carry our descendants into realms of being and experience far beyond our present imaginings. It is inspiring to reflect that in the deep past, evolution has learned to design her children to live communally, to share and to love, and when our number is up, to sacrifice our very lives that the community might continue to change and to evolve.
-------------------------------------------
Joshua Mitteldorf is a researcher in the Biology Dept of the University of Pennsylvania. He specializes in evolutionary theory and computer simulations that demonstrate how evolution can work on groups and whole populations. He has taught at Harvard, Berkeley and Bryn Mawr.
Originally an astrophysicist, Mitteldorf moved to evolutionary biology as a primary field in 1996.
In private life, Mitteldorf is a musician and environmentalist, an activist against Big Tobacco and advocate for public health. He is married to Alice Ballard, a civil rights attorney, and their two daughters, Sarah and Madeline, are among the first children adopted from China in the mid-1980s.